Joint Research Group Macromolecular Crystallography
Structure of the month - December 2013
J. Biol. Chem. jbc.M113.510461 (2013).
Structure of the cyanobacterial phytochrome 2 photosensor implies a tryptophan switch for phytochrome signaling
Katrin Andersa, Grazia Daminelli-Widanyb, Maria Andrea Mroginskib, David von Stettenc, Lars-Oliver Essena*
a Biomedical Research Centre / Department of Chemistry, Philipps-Universität, D-35032 Marburg, Germany
b Department of Chemistry, Technische Universität Berlin, D-10623 Berlin, Germany
c European Synchrotron Radiation Facility, F-38043 Grenoble Cedex, France
* Corresponding author: Lars-Oliver Essen, Biomedical Research Center / Department of Chemistry, Philipps-Universität, D-35032 Marburg, Germany
E-mail: essen@chemie.uni-marburg.de
Phone: (+49)6421-28-22032, Fax: (+49)6421-28-22191
Abstract
Phytochromes are highly versatile photoreceptors, which occur ubiquitously in plants as well as in many light-responsive microorganisms. Here, photosynthetic cyanobacteria utilize up to three different phytochrome architectures, where only the plant-like and the single-domain cyanobacteriochromes are structurally characterized so far. Cph2 represents a third group in Synechocystis species and affects their capability of phototaxis by controlling c-di-GMP synthesis and degradation. The 2.6 Å crystal structure of its red/far-red responsive photosensory module in the Pr state reveals a tandem-GAF bidomain that lacks the figure-of-eight knot of the plant/cph1 subfamily. Its covalently attached phycocyanobilin chromophore adopts a highly tilted ZZZssa conformation with a novel set of interactions between its propionates and the GAF1 domain. The tongue-like protrusion from the GAF2 domain interacts with the GAF1-bound chromophore via its conserved PRxSF, WxE, and WG/AG motifs. Mutagenesis showed that the tongue’s integrity is indispensable for Pr→Pfr photoconversion and involves a swap of the motifs’ tryptophans within the tongue-GAF1 interface. This ‘Trp switch’ is supposed to be a crucial element for the photochromicity of all multi-domain phytochromes.
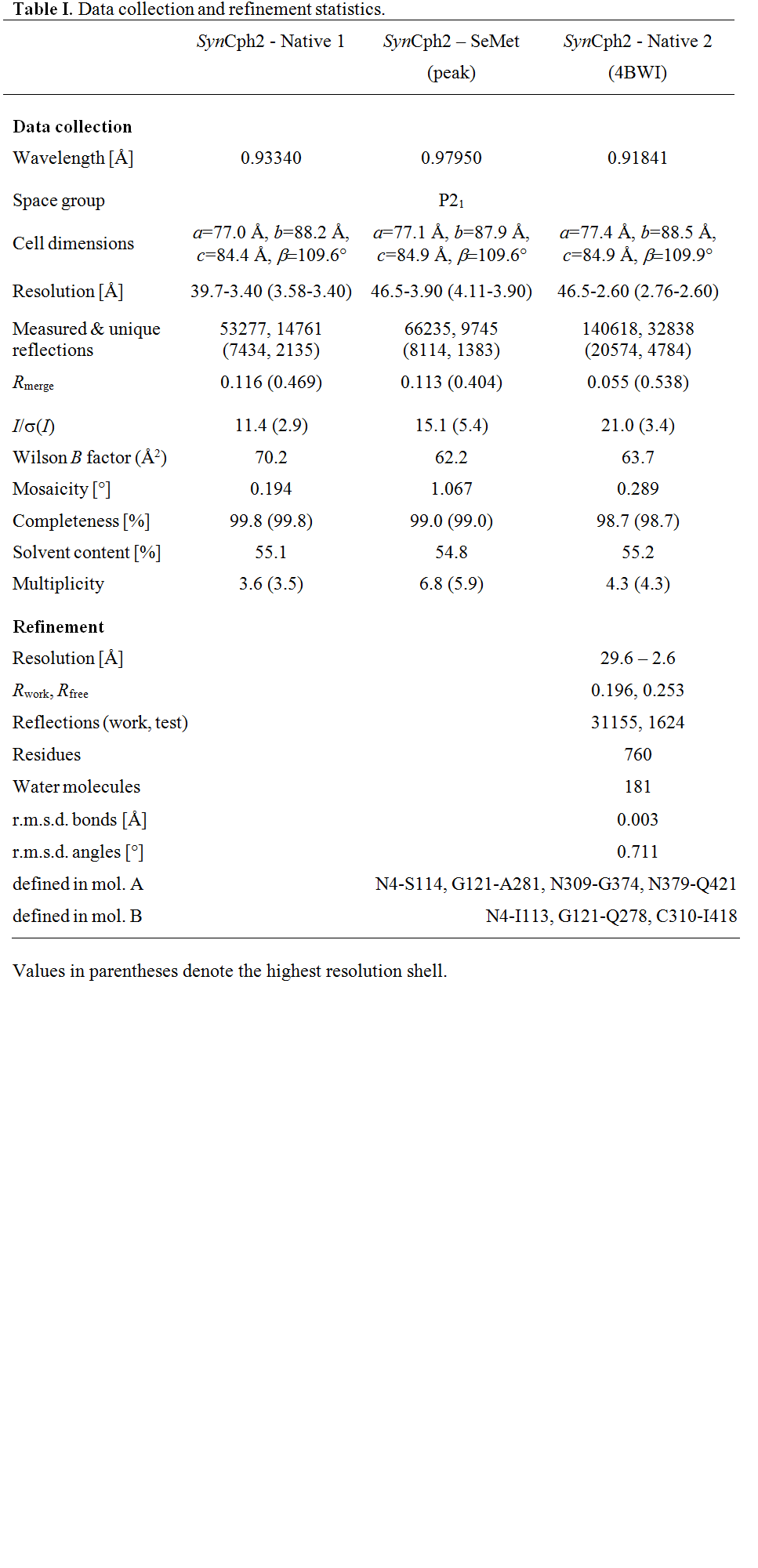
Table 1.

Figure 1.
Structure of the SynCph2(1-2) photosensor. (A) Domain organization of SynCph1 and SynCph2. The tonguelike regions protruding from the GAF2 and PHY domains are shown in green and purple, respectively; the knot in SynCph1 as black lines. SynCph1 and SynCph2 differ in their effector domains: a histidine kinase domain (HKD) and cdiGMP turn-over (GGDEF and EAL) and regulatory (GGDEF*) domains. (B) Crystal structure of the SynCph2(1-2) module. GAF2 (green) contains a tonguelike region (dark-green) for sealing the PCB (cyan)-binding pocket. The GAF1 domain is in orange.
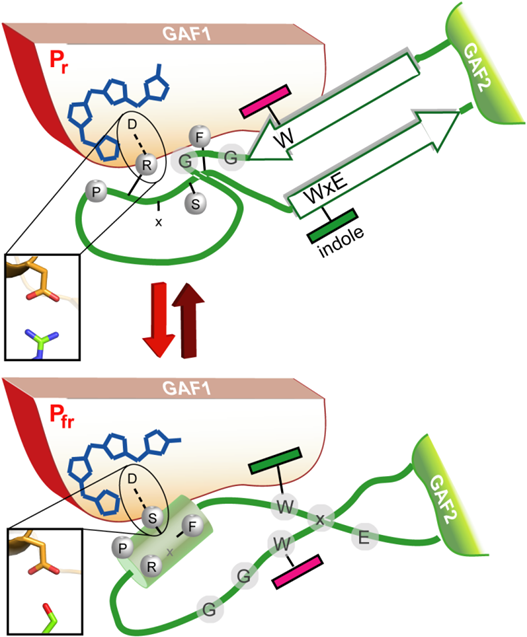
Figure 10.
A model for the Trp-motif switch within phytochromes. Upon red-light triggered formation of the Pfr state and breakage of the tongue/GAF1 Asp-Arg salt bridge (inlet) the tip of the tongue refolds with concomitant disordering of the stalk-like -hairpin and formation of an Asp-Ser salt bridge (inlet).